© Klaus Rajewsky Lab, MDC
K. Rajewsky Lab
Immune Regulation and Cancer
Profile
Studying human B and T cell pathologies genetically, in preclinical mouse models and through gene repair in hematopoietic stem cells and mature lymphocytes
We combine Cre/loxP- and CRISPR/Cas-mediated targeted mutagenesis to study hematopoietic development with an emphasis on B cell physiology and antibody responses; and pathologies such as B cell malignancies, diseases caused or impacted by Epstein-Barr-Virus infection (including Multiple Sclerosis), and primary immunodeficiencies. In the latter contexts we develop preclinical genetic mouse models, and genetically modify human and mouse hematopoietic cells like T and B lymphocytes and hematopoietic stem cells. To this end we have established highly efficient methods of gene repair in such cells, suitable for application in somatic cell therapy.
Free download of recent publications:
Team




Research
B cell development: B cell receptor (BCR) specificity determines B cell subset differentiation
B lymphocytes are the carriers of humoral immunity and develop into two fundamentally different subsets: Classical B2 cells in spleen and lymph nodes, involved in antigen-driven antibody responses, and B1 cells in the peritoneal and pleural cavities, that are responsible for the production of evolutionary selected “natural” antibodies. In experiments that we had started at Harvard Medical School and completed at the MDC, we had shown that the differentiation of B cells into “innate” B1 or “adaptive” B2 cells is determined by the antigen specificity of the BCR expressed by the cells, with BCR autoreactivity driving B1 differentiation. This was shown by genetically switching BCR specificity in mature B2 cells. Although this approach does not directly mimic B1 cell development in normal physiology, it provides a unique opportunity to study B1 subset differentiation in a defined in vitro setting, with B2 cells apparently retaining BCR specificity dependent plasticity with respect to assuming a B1 phenotype. In ongoing work, collaborating with groups at the MDC and the Karolinska Institutet in Stockholm, we explore the mechanism by which, upon BCR specificity switch, B2 cells turn into B1 cells within a 1 – 2 day window, by determining changes of BCR engagement and signaling, as well as gene expression and epigenetic changes over time, determined at the single cell level. These experiments are complemented by attempts to identify key molecular players by genetic CRISPR screens.
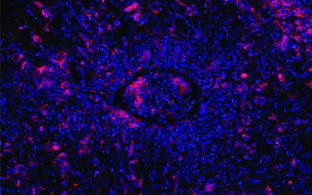
The germinal center reaction, plasma cell differentiation, and B cell malignancies
In the germinal center (GC) reaction antigen-activated B cells proliferate, undergo (early) class switch recombination and somatic hypermutation of their rearranged antibody variable region genes, and are selected for high-affinity antigen binders. Because both class switch recombination and somatic hypermutation involve DNA breaks whose repair can go wrong, the GC reaction is at the origin of most B cell malignancies. Our group has a long tradition in the analysis of these processes and in modeling GC-derived B cell malignancies in mice through conditional gene targeting. With respect to B cell malignancies, work over the recent years and ongoing experiments address the pathogenesis of Burkitt lymphoma (with special emphasis on the roles of PI3K signaling, the transcription factor FOXO1 and miRNAs controlling the Bim-mediated proapoptotic activity of the MYC oncogene), Waldenström’s macroglobulinemia, and, in collaboration with the Janz/Mathas group at the MDC/ECRC, Multiple Myeloma, a malignancy of plasma cells. Multiple Myeloma represents a number of genetically and clinically distinct disease entities, for which we have generated first preclinical genetic mouse models. CRISPR screens of normal plasma cell differentiation are undertaken in parallel.
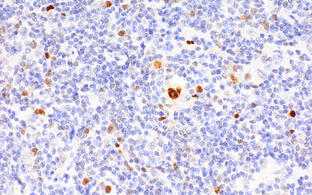
Analyzing Epstein-Barr-Virus (EBV) induced pathologies in mouse models and human B cells
EBV is a B cell transforming herpes virus endemic in humans, with most of the human population latently infected worldwide. Acute infection passes without symptoms in young children and causes infectious mononucleosis (“kissing disease”) in young adults, with initial dramatic expansion of infected B cells and ensuing control by cytotoxic T and NK cells through life-long immune surveillance. EBV then persists latently in a small fraction of B cells. Reactivation under conditions of immunosuppression (such as in post-transplantation patients) and EBV infection in children suffering from hereditary “primary” T and NK cell immunodeficencies can lead to life-threatening lymphoproliferative diseases including B cell lymphomas. In addition, recent studies have identified EBV infection as a precondition for the development of Multiple Sclerosis. We have developed mouse models of EBV acute infection, also modeling surveillance of “infected” cells by T cells, and EBV-induced B cell lymphomagenesis through the conditional expression of EBV oncogenes in mouse B cells. We use these preclinical models together with analyses of human B cells infected and transformed by individual or combinations of such genes for a better understanding of the genetic basis of EBV induced pathologies. In one line of the work, we model the dramatic effect of EBV infection in patients suffering from inherited perforin deficiency, and show that in the mouse model the animals can be cured by transfer of perforin-repaired T cells (see paragraph below). In another line of experiments, we have shown in collaboration with the group of Volker Siffrin at the ECRC that in our model of acute EBV infection mice carrying a transgenic T cell receptor with specificity for the myelin protein MOG tend to develop Experimental Allergic Encephalitis, potentially providing a link between EBV infection and Multiple Sclerosis pathogenesis as observed in humans.
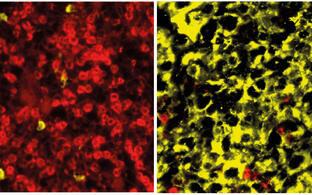
Gene repair in mouse and human T and hematopoietic stem cells
In the course of the work outlined above we have made, in collaboration with Ralf Kühn’s group, substantial efforts to improve CRISPR/Cas mediated targeted mutagenesis and gene repair in cells of the hematopoietic system, particularly in lymphocytes and hematopoietic stem cells of mouse and human. We can now introduce mutations and repair mutant genes in these cells with high efficiency and specificity. In this connection we recently developed a variation of traditional CRISPR/Cas mutagenesis that uses a Cas9 nickase and sgRNAs targeting base sequences on opposite strands of the target gene at a distance (“spacer-nick”). This largely avoids both off- and on-target errors of repair through non-homologous end joining, preserving high rates of homologous repair through recombination with a donor template provided to the cells by a viral or other vector (collaboration with the MDC groups of Kathrin de la Rosa and Ralf Kühn). We are using CRISPR/Cas-mediated targeted mutagenesis in a variety of contexts including some of the work described in the sections above. Thus, we have shown in our mouse model that we can cure the severe disease arising under the condition of perforin deficiency upon EBV infection, and that we can efficiently repair mutations like those observed in perforin and other life-threatening immune deficiencies (affecting e. g. neutrophil elastase in the human) in both human and mouse T and hematopoietic stem cells. In general terms, CRISPR/Cas-mediated mutagenesis in hematopoietic stem cells, with subsequent somatic reconstitution of the hematopoietic system, provides a unique opportunity to generate mouse models, also humanized mouse models, within an unprecedented short time frame. It also brings us close to medical somatic gene therapy.
Selected recent references
Tran, N.T., Danner, E., Li, X., Graf, R., Lebedin, M., de la Rosa, K., Kühn, R., Rajewsky, K., Chu, V.T. (2022). Precise CRISPR-Cas-mediated gene repair with minimal off-target and unintended on-target mutations in human hematopoietic stem cells. Sci Adv 8(22):eabm9106.
Winkler, W., Farré Díaz, C., Blanc, E., Napieczynska, H., Langner, P., Werner, M., Walter, B., Wollert-Wulf, B., Yasuda, T., Heuser, A., Beule, D., Mathas, S., Anagnostopoulos, I., Rosenwald, A., Rajewsky, K., Janz, M. (2021). Preclinical models of human multiple myeloma subgroups. bioRxiv 2021.08.28.458010
Tran, N.T., Graf, R., Wulf-Goldenberg, A., Stecklum, M., Strauß, G., Kühn, R., Kocks, C., Rajewsky, K., Chu, V.T. (2020). CRISPR/Cas9-mediated ELANE mutation correction in hematopoietic stem and progenitor cells to treat severe congenital neutropenia. Molecular Therapy 28, 2621 – 2634.
Schmidt, K., Sack, U., Graf, R., Winkler, W., Popp, O., Mertins, P., Sommermann, T., Kocks, C., Rajewsky, K. (2020). B‑cell-specific Myd88 L252P expression causes a premalignant gammopathy resembling IgM MGUS. Frontiers in Immunology 11, 602868.
Sommermann, T., Yasuda, T., Ronen, J., Wirtz, T., Weber, T., Sack, U., Caeser, R., Zhang, J., Li, X., Chu, V.T., Jauch, A., Unger, K., Hodson, D.J., Akalin, A., Rajewsky, K. (2020). Functional interplay of Epstein-Barr virus oncoproteins in a mouse model of B cell lymphomagenesis. Proc Natl Acad Sci U S A 117, 14421 – 14432.
Graf, R., Seagal, J., Otipoby, K.L., Lam, K.P., Ayoub, S., Zhang, B., Sander, S., Chu, V.T., Rajewsky, K. (2019). BCR-dependent lineage plasticity in mature B cells. Science 363, 748 – 753.
Wong, J.B., Hewitt, S.L., Heltemes-Harris, L.M., Mandal, M., Johnson, K., Rajewsky, K., Koralov, S.B., Clark, M.R., Farrar, M.A., Skok, J.A. (2019). B‑1a cells acquire their unique characteristics by bypassing the pre-BCR selection stage. Nature Communications 10, 4768.
Labi, V., Peng, S., Klironomos, F., Munschauer, M., Kastelic, N., Chakraborty, T., Schoeler, K., Derudder, E., Martella, M., Mastrobuoni, G., Hernandez-Miranda, L.R., Lahmann, I., Kocks, C., Birchmeier, C., Kempa, S., Quintanilla-Martinez de Fend, L., Landthaler, M., Rajewsky, N., Rajewsky, K. (2019). Context-specific regulation of cell survival by a miRNA-controlled BIM rheostat. Genes & Development 33, 23 – 24.
Kabrani, E., Chu, V.T., Tasouri, E., Sommermann, T., Baßler, K., Ulas, T., Zenz, T., Bullinger, L., Schultze, J.L., Rajewsky, K., Sander, S. (2018). Nuclear FOXO1 promotes lymphomagenesis in germinal center B cells. Blood 132, 2670 – 2683.