
Zelluläre und intrazelluläre Bewegungsmechanismen
Viele Zellen sind in der Lage, sich auf einer festen Oberfläche fortzubewegen. Die für die Bewegung notwendigen Kräfte werden bei vielen Zelltypen durch ein intrazelluläres Netzwerk aus Aktinfilamenten erzeugt (Pollard, 2003). Dies erfolgt nach dem folgenden physikalischen Prinzip: Die Membran schränkt die Brownsche Bewegung der semi-flexiblen Aktinfilamente ein, deren freie Energie dadurch erhöht wird. Die Zunahme der freien Energie eines einzelnen Filaments hängt dabei stark vom Abstand des Filamentursprungs zur Membran ab. Dadurch spüren sowohl Membran als auch das Aktinfilament eine sogenannte entropische Kraft, die sich aus der Ableitung der freien Energie nach der Positionskoordinate ergibt. Diese Kraft kann mit Standardmethoden der statistischen Physik berechnet werden (Gholami, 2006) und liefert die Basis für die mathematische Modellierung des darauf basierenden Bewegungsmechanismus.
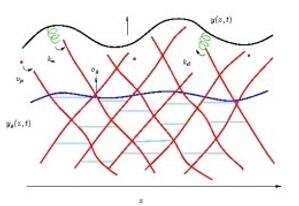

Um die globale Bewegung der Membran zu modellieren, müssen in jedem Punkt der Membran die durch das gesamte Aktinnetzwerk ausgeübten Kräfte berechnet werden. Dafür müssen neben der Längen- und Orientierungsverteilung der Filamente, die explizit in den Kraftausdruck eingehen, auch verschiedene intrazelluläre Prozesse berücksichtigt werden: Filamente können an die Membran binden und wieder los lassen, die frei fluktuierende Länge ändert sich infolge von Polymerisation (Längenwachstum) und Gelbildung (Verkürzung), neue Filamente können durch Verästelung alter Filamente entstehen, das Wachstum polymerisierender Filamente kann von gewissen Proteinen gestoppt werden (Kappung).
Die mathematische Modellierung all dieser intrazellulären Prozesse führt zu Systemen von nichtlinearen partiellen Differentialgleichungen. Diese können analytisch untersucht (explizite Berechnung und Stabilittsuntersuchung von Lösungen mit gewissen Symmetrien, zum Beispiel stationäre Lösungen) und numerisch simuliert werden. Auf diese Weise ist es möglich, die Geschwindigkeit und Membranmorphologie der Zelle zu bestimmen und auch den Einfluss einiger Parameter (Monomerkonzentration, Bindungsrate, Kappungsrate usw.) auf die Zellbewegung zu studieren. Da diese Parameter durch die Zelleigenschaften festgelegt und nur schwer experimentell veränderbar sind, ist die Rolle der Theorie bei dem Verständnis der Kontrollmechanismen zellulärer Motilitt entscheidend.
Weiterhin gibt es eine Reihe von Modellsystemen, in welchen Teile dieses komplexen Bewegungsmechnismus separat verstanden werden können: Einige Bakterien nutzen das Aktinnetzwerk der befallenen Zelle, um sich selbst durch die Zelle zu bewegen und von Zelle zu Zelle zu wandern. Sie bleiben dabei ausserhalb der Reichweite der körpereigenen Immunabwehr und können so immer mehr Zellen befallen (Hamon, 2006). Es ist weiterhin möglich, die Bewegung dieser Bakterien in vitro mit Hilfe von beschichteten Plastikkugeln oder ltropfen zu rekonstruieren (Bernheim-Groswasser, 2005). Dabei lassen sich auch experimentell verschiedene Parameter variieren, wie zum Beispiel Kugelradius, oder Zusammensetzung der Lösung. Diese Experimente können genutzt werden, um die Vorhersagen der Theorie im Einzelnen zu testen und einige Modellparameter anzupassen.
Das theoretische Verständnis von zellulären und intrazellulären Bewegungsmechanismen ist schließlich auch deshalb sehr wichtig, weil eine Reihe von Körperfunktionen darauf basieren: Während der Morphogenese bewegen sich Zellen zielgerichtet zu der Stelle hin, wo sie spezifische Gewebe und Organe bilden; Zellen des Immunsystems müssen sich zu der infizierten Stelle hinbewegen, um die Infektion verursachenden Mikroorganismen zu beseitigen; damit eine Wunde heilt, bewegen sich darauf spezialisierte Zellen zu der Wundstelle hin, um die zerstörten Zellen zu ersetzen und das Gewebe zu remodellieren. Obwohl sie strukturell und funktionell verschieden sind, scheinen all diese Zellen einen universellen Bewegungsmechanismus zu nutzen, das auf Aktinpolymerisation basiert (Ananthakrishnan, 2007).